Электропроводность живых тканей
Электрические свойства биологических тканей.
При решении основных медицинских задач диагностики и терапии заболеваний широко используются электромагнитные явления, что, в свою очередь, предполагает знание электромагнитных характеристик биологических тканей.
Современные представления об электрических и магнитных свойствах живых тканей основаны на фактах о молекулярной организации биологических мембран и, в значительно меньшей степени, на сведениях о квантово‑механических свойствах физиологически активных молекул. Характеризуя электрические свойства живых тканей, следует учитывать, что они являются композиционными средами, т.е. одни структурные элементы обладают свойствами проводников, а другие — диэлектриков, поэтому основное внимание при исследованиях уделяется электропроводящим и диэлектрическим свойствам живых тканей.
Упорядоченное движение свободных зарядов, возникающее в проводнике под действием электрического поля, называется током проводимости. Сила тока равна заряду, протекающему по проводнику за одну секунду, обозначается I и измеряется в амперах (А).
Закон Ома связывает силу тока (I), электрическое напряжение (U) и сопротивление проводника (R): I=U/R. Величина обратная сопротивлению (1/R) есть проводимость проводника, обозначается G и измеряется в сименсах (См= Ом ‑1 ).
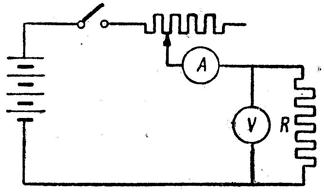
Рис.5. Схема измерения проводимости или сопротивления.
Электропроводность целых органов на 4–6 порядков ниже электропроводности жидкостей, находящихся в них. Это связано с тем, что электролиты составляют малую часть клетки. В клетках электролиты заключены в мельчайшие отсеки — «компартменты», образованные биологическими мембранами, которые составляют более 50% массы клетки. Мембраны же являются диэлектриками.
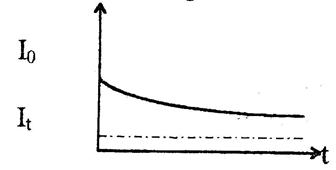
Рис. 6. Зависимость силы тока, текущего через живую ткань при постоянном приложенном напряжении, от времени.
Процесс измерения проводимости связан с определением силы тока, текущего через проводник, измерения напряжения, приложенного к проводнику, и расчету сопротивления по закону Ома. Схема измерения показана на рис. 5. Однако измерение проводимости биологических тканей на постоянном токе невозможно из‑за больших погрешностей, связанных с зависимостью силы тока от времени измерения. С течением времени ток уменьшается до определенного уровня. Эта зависимость представлена на рис. 6. Релаксационный процесс установления равновесия можно объяснить поляризацией живой ткани. Поляризация — это процесс перемещения связанных зарядов под действием электрического поля и образования вследствие этого электродвижущей силы, которая направлена противоположно внешнему полю. Ее называют ЭДС поляризации. Для живой ткани закон Ома можно записать в виде: I=U-Eп/R, где U — приложенное напряжение, I — сила тока, R — активное сопротивление ткани, Eп — ЭДС поляризации, зависящая от приложенного напряжения и времени. Изменение силы тока может быть обусловлено и процессом заряда тканевых емкостей. Релаксационный процесс не позволяет однозначно измерить проводимость живой ткани.
Чтобы релаксационный процесс не влиял на результат измерения проводимости живых тканей, необходимо использовать переменные токи. Простейший переменный ток — синусоидальный, у которого только два из трех независимых параметров — амплитуда и частота. Полное сопротивление электрической цепи при синусоидальных напряжениях и токах называют импедансом и обозначают Z. В общем случае, импеданс имеет активную R и реактивную компоненту, связанную с сопротивлением конденсатора или индуктивности. Сопротивление конденсатора RС=1/ωС, сопротивление индуктивности RL=Lω, активное сопротивление R от частоты не зависит. Клеточные мембраны биологических клеток определяют емкостное сопротивление.
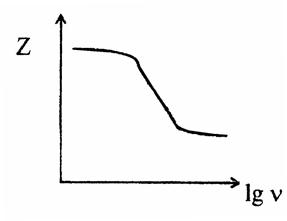
Рис. 7. Схематичная зависимость импеданса живой ткани от частоты протекающего через неё электрического тока.
Омическое сопротивление зависит от ионной проводимости. Индуктивное сопротивление биологической ткани крайне мало (Glaser), поэтому можно считать, что импеданс живой ткани содержит только активную и емкостную составляющие. При определении импеданса электрической цепи, включающей резисторы и конденсаторы, необходимо также учитывать сдвиг фазы, отражающий свойства диэлектриков. Для биологических объектов характерен большой сдвиг фаз между силой тока и напряжением, что говорит о значительной доли емкостной составляющей в полном сопротивлении. Для кожи человека, например, при частоте 1 кГц сдвиг фаз равен 55°.
Для живых тканей характерно уменьшение импеданса по мере повышения частоты внешнего электромагнитного поля. Зависимость импеданса от частоты называют дисперсией импеданса (рис. 7). Импеданс живой ткани зависит от ее физиологического состояния, и его значение может быть использовано для диагностики. Диагностический метод, основанный на измерении импеданса тканей, называется реографией.


Рис. 8. Электрические модели живой ткани.
В клинической практике используются следующие основные методы оценки электрических параметров живых тканей.
1. По кривой Z(ω) оценивают уровень обмена веществ и жизнеспособность органов и тканей, определяя коэффициент поляризации Кп=Zнч/Zвч.
(Zнч=10 2 Гц; Zвч=10 6 Гц). Жизнеспособная ткань имеет Кп>1, причем значения коэффициента поляризации тем больше, чем выше уровень обмена веществ в ткани и чем лучше сохранена ее структурная целостность. При отмирании ткани ее коэффициент поляризации стремится к 1. Этот метод используется для оценки жизнеспособности тканевых трансплантатов при пересадке органов, для определения зоны раневого процесса в ходе хирургической обработки ран, для характеристики ишемии, отека и т.д., и т.п.
2. Метод реоплетизмографии позволяет, изучая динамику изменения активной составляющей импеданса R, судить о кровенаполнении исследуемого органа. Чем больше крови содержится в органе, тем се его электрическое сопротивление.
3. По динамике электрического сопротивления кожи судят о так называемых кожно‑гальванических реакциях, по которым изучают эмоции, утомление и другие состояния организма.
Импеданс живой ткани можно моделировать с помощью эквивалентных схем. На рис. 8приведены две такие схемы.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Живая ткань как проводник переменного электрического тока. Дисперсия электропроводности и её количественная оценка




![]()
Экспериментально установлены следующие особенности живой ткани как проводника переменного ток:
1. Сопротивление живой ткани переменном току меньше, чем постоянному.
2. Электрические характеристики ткани зависят как от её вида, так и от частоты тока.
3. С увеличением частоты полное сопротивление живой ткани нелинейно уменьшается до определенного значения, а затем остаётся практически постоянным (в большинстве на частотах свыше 10 6 Гц)
4. На определенной частоте полное сопротивление зависит также от физиологического состояния (кровенаполнения), что используется на практике. Исследование периферического кровообращения на основе измерения электрического сопротивления называются реография (импедансплетизмография).
5. При умирании живой ткани её сопротивление уменьшается и от частоты не зависит.
6. При прохождении переменного тока через живые ткани наблюдается явление, которое называется дисперсией электропроводности.
Частотные зависимости по форме кривой для разных тканей сходный, но отличается значением сопротивления.
Особенности дисперсии:
1. Присуща только живым тканям.
2. Более выражена на частотах до 1 МГц.
3. На практике используется для оценки физиологического состояния и жизнеспособности тканей.
Количественно оценка дисперсии проводиться по коэффициенту дисперсии (К).
Коэффициент дисперсии это безразмерная величина, равная отношению низкочастотного (10 2 ) полного (или удельного) сопротивления к высокочастотному (10 6 Гц).
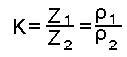
Z1 – полное сопротивление на частоте 10 2 Гц
Z2 – полное сопротивление на частоте 10 6 Гц
Явление дисперсии связывают с наличием в живых тканях поляризации, которая с увеличением частоты меньше влияет на полное сопротивление. Поэтому коэффициент дисперсии часто называют коэффициентом поляризации.
Кроме частотных зависимостей в живых тканях отмечаются фазовые сдвиги между током и напряжением, которые тоже, но в меньшей степени, зависят от частоты.
Фазовые сдвиги тоже уменьшаются при умирании тканей и, в перспективе, могут быть использованы для практических целей.

Дисперсия диэлектрической проницаемости объясняет возможность селективного нагревания разных тканей в высокочастотных ЭМП. Возрастание интенсивности движения ионов, переориентации полярных молекул в ЭМП связаны с поглощением энергий поля и аналогичны процессу нагревания.
Для последовательного соединения этих элементов импеданс равен: 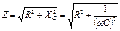
Для параллельного соединения активного и емкостного сопротивлений: 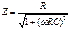
(электроемкость плоского конденсатора 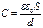 )
)
Электроемкость живых тканей складывается из емкости компартментов, так как клеточные мембраны можно считать диэлектриками, а клеточные и межклеточные жидкости, содержащие растворы солей, проводниками. В момент включения тока конденсаторы заряжаются и величину электроёмкости ткани можно оценить по формуле:
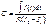
Электрические свойства живой ткани можно смоделировать следующими цепями:

Зависимость импеданса живой ткани (а) и её моделей (б) от частоты тока представлена на рис.

Отметим, что ход кривых дисперсий Z и ε в одной и той же ткани несовпадает. Дисперсия Z, очевидно, отражает более широкий круг электрических процессов в живых тканях. Импеданс сильнее зависит от разнообразньх нарушений жизнедеятельности исследуемой ткани. Есть упрощенный, но эффективный способ определения жизнеспособности ткани. Определяют коэффициент поляризации:
В живой ткани К>1 и тем больше, чем выше уровень обмена веществ и чем лучше сохранена структурная целостность.
Широкое применение в медицинской практике нашла методика реографии— исследование импеданса тканей, органов, обусловленных изменением их кровенаполнения.
В этом методе используют обычно ток частотой от 20 до 200 кГц, и не более 10 мА в величине. Реограмма представляет собой зависимость импеданса ткани от времени. Конфигурация, амплитуда и временные параметры этой кривой зависят от многочисленных факторов: состояния миокарда, тонуса сосудов, состояния капиллярного и венозного кровотока.
В процессе жизнедеятельности живые организмы, человек в частности, излучают в широком диапазоне электромагнитных волн: от радиочастот до ультрафиолетового излучения. В широком диапазоне частот излучаются и звуки. Тело человека имеет сложное строение, т.е. подразделяется на отдельные органы, ткани, жидкости; оно характеризуется значительной неоднородностью пассивных электрических, механических и др. характеристик. Все это обуславливает сложную структуру излучаемых ЭМ и звуковых полей. К тому же все излучения обязательно промодулированы переменными физиологическими процессами.
Физические поля несут обширную информацию о жизнедеятельности живых организмов. Поэтому понятен интерес к их регистрации, изучению и применению в диагностике. Рассмотрим более подробно поля излучаемые человеком.
1. Для тела с температурой около 300 К максимум излучения приходится на инфракрасную область с λ=8-14 мкм. Мощность излучения человека в этой области составляет около 100 Вт.
Злокачественные опухолевые процессы повышают температуру тканей на 1,2°С, при нарушениях кровообмена температура в органе снижается.
Увеличение глубины, на которой измеряется температура, возможнo при увеличении длины волны регистрируемого излучения, но при этом ухудшается разрешающая способность в определении изменений температуры.
Исследование низкочастотных полей очень распространено в диагностике. Это такие методы как электрокардиография, электроэнцефалография, электромиография.
4.Излучение магнитных полей.
Основные химические компоненты биосред относятся к диамагнетикам (белки, углеводы, липиды, вода) с магнитной проницаемостью µ≈1, это означает, что живые ткани не ослабляют магнитное поле.
Источники магнитных полей можно локализировать с достаточной точностью, несмотря на то, что человек излучает очень слабые магнитные поля, примерно, в миллион раз слабее магнитного поля Земли. Магнитодиагностика весьма информативна в исследованиях сердца, мозга, мышц. Так, при исследовании сердца магнитные сигналы позволяют локализовать область патологии с точностью до 1 см.
5. В оптическом диапазоне регистрируются сигналы биолюминесценции, по которым можно контролировать протекание биохимических процессов.
6.В процессе метаболизма биологический объект вносит возмущение в среду, т.е. изменяет газовый и аэрозольный состав, проводимость и диэлектрическую проницаемость. Например, по запаху можно диагностировать около 40 заболеваний: абцесс легкого, диабет, дифтерию и др.
7.Очень информативны акустические сигналы. В инфразвуковом диапазоне может быть получена информация о функционировании внутренних органов, мышц. О процессах на клеточном и молекулярном уровне несут информацию высокочастотные сигналы.
Источники акустического излучения можно локализовать достаточно точно, т.к. длина акустических волн меньше, чем у электромагнитных (низкочастотных) волн.
Состояние биоо6ъекта существенно не стационарно, поэтому важно наблюдать динамику изменения физических полей, в которой должны проявляться характеристики регуляторных систем гомеостаза.
Впервые термин «биополе» или «морфогенное поле» ввел в научный оборот А.Г. Гурвич. В результате экспериментов по морфогенезу он пришел к следующим выводам:
1.Вокруг растущего зародыша существует некое «морфогенное поле», распространяющееся за его пределы и определяющее в каждой точке пространства направление и скорость роста других клеток того же организма.
2.Морфогенные поля отдельных зародышевых клеток векторно складываются.
3.Морфогенное поле имеет не силовой характер, т.е. само не совершает никакой работы против внешних сил. Оно выполняет сугубо информационную роль, управляя процессами обмена веществ в клетках.
Мы видим, что введенное понятие является полем лишь с точки зрения биолога, но не физика. Позднее этим термином стали называть «новое» физическое поле, а это уже подпадает под.
Чтобы распечатать файл, скачайте его (в формате Word).
Электропроводность в неживой природе. Электропроводность живых тканей. Диэлектрические свойства живых тканей. Физические поля биологических объектов
Страницы работы

Содержание работы
ПАССИВНЫЕ ЭЛЕКТРИЧЕСКИЕ СВОЙСТВА ЖИВЫХ ТКАНЕЙ.
Основными пассивными электрическими характеристиками вещества являются: диэлектрическая проницаемость, удельная электрическая проводимость. Эти величины и связанные с ними электроемкость, импеданс существенно зависят от состояния организма и активно используются в диагностике
1) ЭЛЕКТРОПРОВОДНОСТЬ В НЕЖИВОЙ ПРИРОДЕ
Пассивные электрические свойства, веществ в неживой природе хорошо изучены. Переходя к изучению свойств живых тканей (они значительно сложнее) мы будем опираться на их неживые модели.
Электропроводность — это величина, обратная электрическому сопротивлению G=1/R
Удельная электрическая проводимость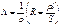
Закон Ома можно переписать через плотность тока, удельную электрическую проводимость и напряженность электрического поля
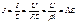
Рассмотрим электропроводность в неживой природе. В зависимости от вида и природы носителей проводимость бывает электронной, ионной и дырочной. Электронной проводимостью обладают металлы. Жидкие электролиты обладают ионной проводимостью. Дырочная проводимость образуется при разрыве валентной связи, приводящей к появлению вакантного места для электрона. Полупроводники обладают электронной и дырочной проводимостью.
Электрическая проводимость прямо и косвенно зависит от систем атомов и молекул. Она обусловлена положением электронов относительно ядра, их спинами и магнитными моментами. В металлах некоторые электроны утрачивают связь со своими атомами и могут свободно передвигаться. В растворах основными носителями зарядов являются ионы. Прохождение тока через растворы вызывает перемещение вещества в нем, и. следовательно, изменение его химических свойств. Этим растворы отличаются от металлических проводников.
Современные представления об электрических свойствах живых тканей основаны на фактах о молекулярной организации мембран и, в значительно меньшей степени, на сведениях о квантовомеханических свойствах физиологически активных молекул.
Живые ткани являются композиционными средами, т.к. одни структурные элементы являются проводниками, а другие диэлектриками. Кроме того, некоторые макромолекулы обнаруживают полупроводниковые свойства (А.Сцент-Дъердьи).
Найдены элементы и даже молекулы, играющие роль доноров и акцепторов, + крайне малые значения энергии связи электронов в биологических молекулах.
Для диагностики важно отметить, что наблюдается различие в проводимости опухолей и нормальной ткани, а так же зависимость проводимости от насыщенности тканей кислородом.
1). При измерении пассивных электрических характеристик живых тканей через них необходимо пропускать электрический ток. что приводит к раздражению тканей и изменению их свойств. Результаты измерений зависят от силы тока, его типа, плотности тока на электродах.
При прохождении постоянного тока через ткани его первоначальная сила не остается постоянной, а падает с течением времени до определенного уровня (рис.).

Это означает, что происходит уменьшение проводимости живой ткани. Процесс установления равновесия (релаксационный процесс) имеет сложную природу, можно предложить лишь некоторые его модели:
Поляризация — это процесс перемещения связанных зарядов под действием электрического поля и образование вследствие этого электродвижущей силы, которая направлена противоположно внешнему полю. Ее называют ЭДС поляризации. Для живой ткани закон Ома можно записать в виде: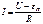
б) процесс заряда тканевых емкостей.
2). Измерения проводимости в БАТ (метод Фолля). Оставляя в стороне достоверность этой диагностики, рассмотрим возможные причины неоднозначности подобных измерений:
а) Исследования проводятся по точкам, расположенным в направлении «меридианов», которые были интуитивно установлены в восточной медицине. Недавно было обнаружено, что в этих направлениях между клетками тканей имеется повышенное число щелевых контактов, проницаемых для ионов, чем и объясняется большая проводимость.
б) При измерениях используют маленькие по площади электроды, и от величины силы, с которой электрод прижимается к коже, существенно зависят получаемые результаты.
в) Ток от внешнего источника так же изменяет состояние БАТ.
г) проводимость меридианов зависит не только от патологий соответствующих органов, но и от индивидуальных биоритмов человека.
3) ДИЭЛЕКТРИЧЕСКИЕ СВОЙСТВА ЖИВЫХ ТКАНЕЙ.
Диэлектрические свойства биологических тканей обусловлены явлениями поляризации и компартментной структурой.
Электрическое поле внутри тканей меньше внешнего так как поляризованные и ориентированные определенным образом атомы, молекулы и компартменты создают электрическое поле Еп направленное против внешнего поля (рис.).
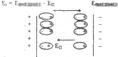
Рассмотрим виды поляризации характерные дляживой ткани.

В живых тканях под воздействием ЭМП возникают токи проводимости, обусловленные смещением (колебаниями) ионов и токи смещения, связанные с переориентацией дипольных моментов. Можно считать что в цепи переменного тока через конденсатор течет именно ток смещения, т.к. перемещения зарядов через конденсатор не происходит.
Электрические свойства биологических тканей
Первое упоминание об исследовании электрической проводимости биологических объектов принято относить к работам В. Томсона, датированным 1880 г. Вильям Томсон (W. Thomson, 1824-1907), с 1892 г. Лорд Кельвин – английский физик, президент Лондонского королевского общества (1890-1895). Основополагающие результаты в исследовании электропроводности биологических тканей были получены в начале и середине ХХ в. Установление типичных значений удельного сопротивления и диэлектрической проницаемости тканей, органов и жидких сред живого организма, а также выявление и частичное объяснение зависимости проводимости и диэлектрической проницаемости биологических жидкостей и клеточных суспензий от частоты зондирующего тока связаны с именами Г. Фрике, К.Коула, Х. Шванна.
Электропроводность биологической ткани – это количественная характеристика способности биомембран, клеток и тканей проводить электрический ток.
Биологические ткани являются композиционными средами, поскольку одни структурные элементы обладают свойствами проводников а другие – диэлектриков. При анализе действия ЭМП на живой организм его ткани принято рассматривать как среду, образованную проводниками и диэлектриками.
 Таким образом, особенность биологических тканей состоит в том, что они обладают свойствами проводников и диэлектриков. Наличие свободных ионов в клетках и тканях обусловливает проводимость. Диэлектрические свойства и величина диэлектрической проницаемости e определяются структурными компонентами биологических тканей и явлениями поляризации. Характерная особенность биологических тканей, как многокомпонентных сложноструктурированных сред – наличие высокой поляризационной ёмкости, высокой диэлектрической проницаемости на низких частотах внешнего воздействующего ЭМП.
Таким образом, особенность биологических тканей состоит в том, что они обладают свойствами проводников и диэлектриков. Наличие свободных ионов в клетках и тканях обусловливает проводимость. Диэлектрические свойства и величина диэлектрической проницаемости e определяются структурными компонентами биологических тканей и явлениями поляризации. Характерная особенность биологических тканей, как многокомпонентных сложноструктурированных сред – наличие высокой поляризационной ёмкости, высокой диэлектрической проницаемости на низких частотах внешнего воздействующего ЭМП.
При постоянном зондирующем токе закон Ома для биологической ткани (рис. 1), как проводника электрического тока, можно записать в виде I(t)=(U—E(t))/R, где U – приложенное напряжение, E(t) – возникающая в ткани противо-ЭДС, R – эквивалентное активное сопротивление цепи «электрод-биообъект-электрод». Уменьшение во времени тока через биоткань обусловлено явлениями поляризации. При прохождении постоянного тока через биологическую ткань в ней возникает нарастающая до некоторого предела ЭДС противоположного направления, которая уменьшает приложенную к ткани эффективную ЭДС, что приводит к уменьшению тока.
Характеристикой поляризации является время релаксации t, соответствующее максимальной частоте внешнего поля, которую индуцированные диполи способны «отслеживать» своими поворотами на 180°, «противодействуя» внешнему полю. Выделяют несколько видов поляризации, возникающих во внешнем ЭМП:
Для импедансных измерений при исследовании биологических тканей наименее существенную роль играют электронная и ионная виды поляризации.
 ,
,
где I0 – начальное значение тока; IT – установившееся через время T значение тока (рис. 1).
При прохождении переменного тока электрические свойства биологических тканей описываются не только активным, но и емкостным сопротивлением.
Все явления поляризации могут быть описаны с помощью диэлектрической проницаемости вещества. Диэлектрическая проницаемость e характеризует уменьшение величины электрического поля в веществе по сравнению с величиной электрического поля в вакууме. Если Е0 – напряженность однородного поля, образуемого некоторыми зарядами в вакууме, а Е – напряжённость поля, создаваемого этими же зарядами, в веществе, то
Если, например, диэлектрическая проницаемость воды равняется 80, то это значит, что напряжённость поля между двумя пластинами, между которыми находится вода, в 80 раз меньше напряженности поля, создаваемого этими же зарядами в вакууме. И это уменьшение напряженности поля обусловлено дипольной поляризацией, вызванной ориентацией (вращением) полярных молекул воды в электрическом поле.
Величина определяется e также как отношение ёмкости С конденсатора, между обкладками которого находится данное вещество, к ёмкости С0 того же конденсатора в вакууме:
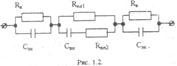
Электрическая модель биологического объекта (исследуемого образца биологической ткани) может быть представлена упрощённо в виде комбинаций поляризационных ёмкостей (C) и активных сопротивлений (R): последовательной (рис. 2, а) и параллельной (рис. 2, б) схемами замещения 3.
 Реактивное ёмкостное сопротивление определяется по формуле
Реактивное ёмкостное сопротивление определяется по формуле
 ,
,
Электрическое сопротивление образца биоткани (или между какими-либо участками тела) представляет собой комплексное сопротивление – импеданс (Z). Импеданс биоткани для последовательного соединения C и R определяется как
 ,
,
 ,
,
где Z – импеданс;  — мнимая единица.
— мнимая единица.
Импеданс биоткани и его составляющие, активная и ёмкостная, изменяются с изменением частоты тока, на котором производится измерение.
Существенное влияние на электропроводность биотканей и биологических объектов (БО) оказывает содержание в них воды. К тканям с низким содержанием воды, около 15% массы ткани, относятся костная и жировая. Их удельная электропроводность невелика: L=0,02 – 0,03 См/м (удельное сопротивление r=1/L=30-50 Ом×м) на частотах ЭМП ниже 1 кГц и около L=0,2 – 0,3 См/м (удельное сопротивление r=3-5 Ом×м) на более высоких частотах, вплоть до 10 ГГц [2].
В целом, электропроводность живых тканей определяется, прежде всего, электрическими свойствами крови, лимфы, ликвора, межклеточной и внутриклеточной жидкостей. Причем подвижность ионов в биологических жидкостях примерно такая же, как и в растворах соответствующих солей, приготовленных на дистиллированной воде.
Живым тканям свойственна зависимость электропроводности от частоты воздействующего ЭМП. Это явление получило название дисперсии электропроводности. С повышением частоты увеличивается и удельная электропроводность. Дисперсия электропроводности установлена для различных биологических тканей в низкочастотном и высокочастотном диапазонах. При изменении частоты ЭМП от 40 Гц до 200 кГц удельная электропроводность мозга кролика увеличивается в 20 раз. На средних частотах дисперсия электропроводности менее выражена, на высоких – проявляется отчетливо. Так, при изменении частоты ЭМП от 25 МГц до 8,5 ГГц сопротивление скелетной мышцы снижается примерно в 10 раз. На 10 ГГц удельная электропроводность жировой ткани достигает 0,3-0,4 См/м (r=2,5 – 3,3 Ом×м), а у костной – 0,5-2,7 См/м (r=0,4 – 2,0 Ом×м) [2].
Дисперсия электропроводности присуща всем неоднородным средам, а не только биологическим. Она наблюдается в том диапазоне частот ЭМП, которые соответствуют характеристическим частотам (nx=1/t) заряженных частиц, входящих в состав той или иной среды. Дисперсия электропроводности – специфическое свойство не только биологических систем, но и других неоднородных сред.
Характерная особенность живых тканей состоит в том, что у них зависимость удельной электропроводности от частоты гораздо отчётливее, чем у сред с менее сложной организацией, и обнаруживается в широком частотном диапазоне. Это обусловлено сложной, мембранной, структурой тканей и большим разнообразием релаксационных способностей их заряженных частиц. Причём такое разнообразие связано как с различиями в размерах, так и с влиянием на их подвижность биологических мембран.
Повреждение клеточных мембран стирает в значительной мере грань между живыми тканями и органическими электролитами в дисперсии электропроводности на низких частотах.
Диэлектрические свойства биотканей определяются присутствием в них малых полярных молекул воды, полярных макромолекул, белков, находящихся в водной фазе, а также компартментализацией клеточных структур.
Компартментализация, обеспечивая оптимальные условия для внутриклеточных биохимических процессов, придаёт живым тканям сегнетоэлектрические свойства.
Каждый органоид (компартмент) на мембране которого поддерживается разность потенциалов между внутри- и внекомпартментным содержимым, имеет значительный дипольный момент и подобен домену в сегнетоэлектрике. За счёт таких заряженных компартментов живые ткани обладают высокой диэлектрической проницаемостью (e). В постоянном электрическом поле она достигает нескольких десятков тысяч.
Как и всякому домену, внутриклеточному компартменту присуща характеристическая частота релаксации (nx). Применительно к диполям nx соответствует максимальной частоте внешнего ЭМП, которую они способны воспроизвести своими полными поворотами в нём, за счёт чего достигается максимально возможная компенсация внешнего поля собственным электрическим полем диэлектрика. Когда диполи уже не успевают совершать повороты на 180° вслед за изменениями направления вектора  ЭМП по мере повышения его частоты, вектор
ЭМП по мере повышения его частоты, вектор  уменьшается по абсолютной величине, что эквивалентно понижению e среды. Области резких изменений e при изменении частоты ЭМП называются областями дисперсии e.
уменьшается по абсолютной величине, что эквивалентно понижению e среды. Области резких изменений e при изменении частоты ЭМП называются областями дисперсии e.
При низких частотах дисперсия e обусловлена, по-видимому, поляризацией внутриклеточных компартментов («доменов»). Диапазон nx разных внутриклеточных компартментов находится в пределах от долей герца до 1 кГц. На более высоких частотах диэлектрические свойства биообъектов определяются полярными молекулами, сосредоточенными как в цитозоле, так и в клеточных мембранах. У разных белковых молекул nx охватывает диапазон от 10 кГц до 100 МГц и зависит от их размеров, а также от вязкости окружающей среды. Эту зависимость выражает следующая формула:
 ,
,
где k – постоянная Больцмана, T – температура среды, в которой вращаются молекулы под действием ЭМП, h – вязкость среды, r – эффективный радиус полярной молекулы.
Важно отметить, что молекулы одинаковых размеров, пребывая в средах с разной вязкостью (например, в цитозоле и в биомембране), обладают неодинаковой nx.
В сверхвысокочастотных полях основной вклад в эти свойства вносит вода, обладающая очень высокой nx. Характеристическая частота релаксации внутриклеточной воды такая же, как и дистиллированной (nx»20ГГц). Именно в воде, входящей в состав живых тканей, происходят основные диэлектрические потери при действии на организм СВЧ-излучений, поскольку с nx воды совпадает частота сантиметровых радиоволн.
 |
Таким образом, поляризация каждого из компонентов живой ткани имеет свою nx. Вблизи неё наблюдается резкое изменение e, которое прекращается, когда частота внешнего ЭМП начинает превосходить nx на порядок.
Неодинаковые величины nx разных тканевых компонентов, способных поляризоваться в ЭМП, обусловливают неравномерный ход кривой дисперсии диэлектрической проницаемости, отображающей зависимость e ткани от n электромагнитных колебаний, воздействующих на неё. На графике рисунка 3 можно выделить три участка, где кривая идёт круче, чем в промежутках. Эти участки называют областями дисперсии (зонами релаксации) и обозначают греческими буквами a, b и g [2, 5].
Первый участок (a-дисперсия живых тканей) соответствует низкочастотному диапазону (до 1кГц). Он отображает поляризацию внутриклеточных компартментов, с которыми связаны сегнетоэлектрические свойства живых тканей. В силу значительной инерционности релаксационных процессов в доменах-компартментах, вращение этих «гигантских диполей» запаздывает относительно перемен направления внешнего ЭМП даже на низких частотах, что проявляется в уменьшении e по мере повышения nx в низкочастотном диапазоне. Некоторый вклад в a-дисперсию вносит релаксация зарядов на фасциях, внутриорганных соединительнотканных прослойках, клеточных поверхностях.
Второй участок (b-дисперсия живых тканей) отображает изменение поляризации макромолекул по мере повышения частоты внешнего ЭМП. В скелетной мышце b-дисперсия наблюдается в диапазоне частот от 10 4 до 10 8 Гц. Снижение e по мере повышения n в этом диапазоне зависит от того, что всё менее крупные макромолекулы не успевают поворачиваться в соответствии с частотой внешнего ЭМП, когда она начинает превосходить nx той или иной полярной молекулы. Очевидно, что 10 8 Гц является частотой, соответствующей nx наименее инерционных макромолекул, а 10 4 Гц – наиболее инерционных макромолекул.
Третий участок (g-дисперсия живых тканей) приходится на n выше 10 10 Гц, чему соответствуют частоты ориентационной поляризации молекул воды. Поскольку воде свойственно несколько значений nx, лежащих около 20ГГц, то изменение e на n > 10 10 Гц имеет немонотонно убывающий характер. Диэлектрическая проницаемость уменьшается потому, что даже такие мелкие молекулы, как H2O, не успевают совершать повороты с частотой, соответствующей частотному диапазону g-дисперсии.
Под действием внешнего ЭМП в живых тканях, обладающих и диэлектрическими, и электропроводящими свойствами, возникают и токи смещения, и токи проводимости. В низкочастотном диапазоне тангенс угла диэлектрических потерь в большинстве тканей равен 20-40, но уже на n = 10 кГц он становится 7 Гц. Сказанное хорошо иллюстрирует различные виды высокочастотной электротерапии: если при диатермии (n = 0,5 – 2 МГц) ткани нагреваются благодаря возникновению в них тока проводимости, то при УВЧ-терапии (n = 40 – 60 МГц) тепловой эффект связан, прежде всего, с током смещения. Можно показать, что при УВЧ-терапии именно ток смещения служит лечебным фактором.
Биоимпедансометрия
В настоящее время в импедансометрии биологических тканей принято рассматривать только две составляющих импеданса (полного сопротивления переменному току) ткани: активное сопротивление R и ёмкостное сопротивление XС 2. Индуктивное сопротивление не учитывается.
Для оценки величины ёмкостного сопротивления, либо оценки величины диэлектрической проницаемости ткани принято использовать упрощённую формулу, определяя С по формуле плоского конденсатора:
 ,
,
где ν – частота внешнего поля в Гц; d – толщина исследуемого образца ткани; s – эффективная площадь сечения образца.
Для биологических тканей характерно уменьшение импеданса с увеличением частоты, т.е. наблюдается частотная дисперсия импеданса [2,5]. Дисперсии L, e и Z не идентичны, но связаны.
Удельное сопротивление биологических тканей, определяемое для заданной частоты тока, может существенно изменяться под влиянием физиологических и патофизиологических факторов: почки и лёгкие изменяют электропроводность при различном крове- и воздухонаполнении, мышечные ткани – при различной степени сокращения мышц, кровь и лимфа – при изменении концентрации форменных элементов, белков и электролитов, очаги повреждения – в результате отёков или ишемий различной природы, опухолей и других причин. Это позволяет использовать биоимпедансометрию для количественной оценки состояния органов и систем организма при различных заболеваниях, а также для выявления изменений в тканях, вызываемых лекарственными, ортостатическими, физическими и другими нагрузками. Однако абсолютные значения импеданса не являются стабильными надёжными показателями, т.к. зависят от многих факторов и не могут рассматриваться как константы живой ткани. Наибольший интерес представляют изменения значений импеданса и его составляющих, а также относительные расчётные параметры.
Существует несколько разновидностей биоимпедансного анализа, которые классифицируются по следующим трём признакам: 1) по частоте зондирующего тока – одночастотные, двухчастотные многочастотные; 2) по объекту измерений – интегральные (объектом измерений служит значительная часть тела), локальные (измеряются отдельные участки тела или регионы), полисегментные (параметры всего организма устанавливаются на основе обработки результатов измерений составляющих его регионов); 3) по тактике измерений – одноразовые, эпизодические, мониторные; 4) по количеству измерительных каналов – одно-, двух- и многоканальные.
Формулы, применяемые в биоимпедансометрии, наряду с характеристиками импеданса, длины исследуемого участка тела расчётные формулы содержат дополнительные параметры, такие как пол, возраст, масса тела, этническая принадлежность, гематокрит и др. для повышения точности оценок (состава тела, ударных объемов крови и т.п.).
Важной характеристикой электрической проводимости тканей является отношение их ёмкостного и активного сопротивлений:  . Величина j в этом уравнении имеет название фазового угла, который характеризует сдвиг фазы переменного тока относительно напряжения [1,5].
. Величина j в этом уравнении имеет название фазового угла, который характеризует сдвиг фазы переменного тока относительно напряжения [1,5].
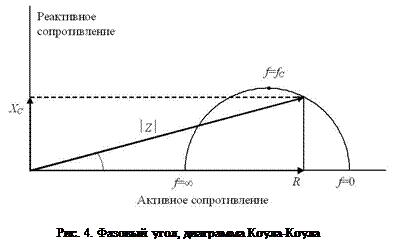 Полуокружность на рис. 4 (график Коула-Коула) описывает теоретическую зависимость между активным и реактивным сопротивлениями тканей при изменении частоты тока f от 0 до +¥. Верхняя точка полуокружности соответствует максимальному значению реактивного сопротивления тканей и характеристической частоте тока. Типичные значения XC и R при измерении импеданса всего тела составляют 20-80 Ом и 200-800 Ом соответственно. Значения j при частоте тока 50 кГц составляют в норме 7,6±1,0° у мужчин и 6,9±1,3° у женщин (пределы изменения от 3 до 10°). При увеличении частоты тока эта величина варьирует в более широких пределах. Пониженные значения XC связывают с нарушением диэлектрических свойств клеточных мембран и увеличением доли разрушенных клеток в тканях организма. Наоборот, повышенные значения ёмкостного сопротивления отражают более высокое функциональное состояние клеточных мембран и, следовательно, самих клеток. На основании этого полагают, что чем больше величина j, тем лучше состояние организма. Повышенные значения XC отражают более высокое содержание активной клеточной массы и трактуются в спортивной медицине как свидетельство тренированности. Николаев Д.В. с соавторами отмечают следующие результаты изучения величины фазового угла для различных популяций: 1) существует высокая корреляция между величиной j и массой тела, у детей с нормальным питанием; дети с пониженным питанием имеют более низкие значения j; 2) величина j снижена у пациентов с почечной недостаточностью (j 6
Полуокружность на рис. 4 (график Коула-Коула) описывает теоретическую зависимость между активным и реактивным сопротивлениями тканей при изменении частоты тока f от 0 до +¥. Верхняя точка полуокружности соответствует максимальному значению реактивного сопротивления тканей и характеристической частоте тока. Типичные значения XC и R при измерении импеданса всего тела составляют 20-80 Ом и 200-800 Ом соответственно. Значения j при частоте тока 50 кГц составляют в норме 7,6±1,0° у мужчин и 6,9±1,3° у женщин (пределы изменения от 3 до 10°). При увеличении частоты тока эта величина варьирует в более широких пределах. Пониженные значения XC связывают с нарушением диэлектрических свойств клеточных мембран и увеличением доли разрушенных клеток в тканях организма. Наоборот, повышенные значения ёмкостного сопротивления отражают более высокое функциональное состояние клеточных мембран и, следовательно, самих клеток. На основании этого полагают, что чем больше величина j, тем лучше состояние организма. Повышенные значения XC отражают более высокое содержание активной клеточной массы и трактуются в спортивной медицине как свидетельство тренированности. Николаев Д.В. с соавторами отмечают следующие результаты изучения величины фазового угла для различных популяций: 1) существует высокая корреляция между величиной j и массой тела, у детей с нормальным питанием; дети с пониженным питанием имеют более низкие значения j; 2) величина j снижена у пациентов с почечной недостаточностью (j 6
10. Записать расчётные данные в таблицу.
11. Построить графики зависимости от частоты зондирующего тока следующих параметров исследуемой мышечной ткани: ZБО(f), R(f), XС(f), C(f), ρ(f) и ε(f).
12. Выполнить аналогичные измерения для образца ткани растительного происхождения (однородного кусочка яблока без кожуры и сердцевины) в соответствии с пп.3 – 7.
13. Рассчитать значения ZБО для яблока на различных частотах, записать полученные значения в таблицу и построить график ZБО(f).
14. Проанализировать полученные результаты и сформулировать выводы по лабораторной работе. Выводы должны отражать следующее: 1) степень соответствия полученных значений и результатов с априорно известной информацией из литературных источников; 2) соответствие анализируемых схем замещения биологической ткани на низких и высоких частотах исследуемого диапазона частот; 3) иметь практическую направленность с точки зрения применения полученных результатов для биомедицинской инженерии.